Since European colonisation begun, exotic species have caused
problems and concerns on biodiversity loss are currently on the
table. We review scientific literature to better understand the role
biodiversity plays on global, regional and local scales and whether
changes in biodiversity composition affect ecosystem function. We
also analyse various Red Lists and reports from world and Brazilian
entities. Species of interest can recover if active predation is
thwarted and conservation efforts undertaken. We conclude that
reports based on Red Lists are exaggerated and there is no threat of
biodiversity loss at sight.
Keywords: biodiversity change, anthropic pressure, community ecology
RESUMO
Desde o começo da colonização europeia, espécies exóticas têm
causado problemas e preocupações sobre perda de biodiversidade
estão em jogo. Revisamos a literatura científica para melhor
entender qual o papel que a biodiversidade executa nas escalas
global, regional e local, e se mudanças na composição de
biodiversidade afetam a funcionalidade do ecossistema. Também
analisamos várias Listas Vermelhas e relatórios de entidades
mundiais e brasileiras. Espécies de interesse podem se recuperar se
predação ativa for barrada e esforços de conservação
empreendidos. Concluímos que relatórios baseados nas Listas
Vermelhas são exagerados e que não há ameaça iminente de perda de
biodiversidade em vista.
Palavras-chave: mudança de biodiversidade, pressão antrópica,
ecologia de comunidades, red list
Preface
Threats
from environmentalist groups about the effects of climate change are
not restricted to the supposed global warming but also include a
pleiad of other misfortunes and extreme events that develop
simultaneously, such as sea level rise, ocean water acidification, and
in the tropical and temperate forests a large loss of biodiversity
with drastic extinction of animal and plant species in an
uncontrolled manner, even menacing survival of the human species. In
this chapter, we propose to debate this prediction under the light of
science, detached from emotional tension and apocalyptic debate that
is propelled by peers in scientific articles and books. Source and
complete research of this paper can be found at the website
<https://biodiversidade.github.io/>.
Introduction
Lamarck, Darwin and, Wallace are
gradualists in their evolution theories. The geologist Niels Eldredge
in the observation of fossils in 1972 noticed long and monotonous
periods without morphological body modifications of species, which
he named ‘stasis’ (from static), and hypothesised that
the same environment pressures directing extinctions are also
responsible for speciation patterns. With that, he formulated the
“punctuated equilibrium” theory in which new speciation patterns
punctuate long and monotonous periods of biological stasis.
Transposable genetic elements (transposons) play a crucial role in
this phenomenon.
Environmental pressure must reach all
the species geographically, and morphological change only occurs after
genetic evolution and when isolated breeding communities can no
longer exchange information with related species (subspecies).
When Darwin was travelling across
South America, Brazil and Argentina, in 1832, in his notes he wondered
what could be the reason for low species diversity of some biological
genera. If space were more important than time in the history of
life, that could explain species stability in an integrated
continental territory such as Brazil over time, in contrast to related
but morphologically distinct species he had observed in archipelagos.
Biological diversity rates can be
calculated subtracting speciation and extinction rates over a
geographical locality (macroecology) or over time (macroevolution).
Since approximately the 1600s, there are well recorded changes in
species composition in various areas in the world.
Excluding archaea, bacteria, fungi, and
virus, that is estimated to be about 1.5 - 1.9 million described species
and a great majority that has not been described yet. Total number of
terrestrial plants must be greater than 450,000. Animal estimates range
from 3 to 11 million species on planet Earth, while insects alone can
range from 5 to 6 million species. Other authors question the possibility
of a plausible estimate when all uncertainties in the number of insects
and fungi are taken under account. Marine species are estimated between
700,000 and 2.2 million, with only approximately 226 thousand described
out of those. Observed average lifetime of invertebrate and vertebrate
species is from one to 10 million years.
In geological history, there were
five great extinction events, all recorded in the Phanerozoic, some
570 million years ago and mostly their
conclusion coincided with the start of speciation events. The
first great speciation event was the Cambrian Explosion of skeletal
marine animals approximately 540 million years ago. The second is
called the Great Ordovician Biodiversification Event (GOBE). That
would be an excessive simplification to think that a dramatic event
unchained big biodiversity pulses in all fossil groups at the global
scale in a particular time frame, for example, GOBE was not a single
event.
There was biodiversity increase in
local richness during Cretaceous (K) and Paleogene (Pg), followed by
relative stasis until present terrestrial tetrapods. Stasis pattern
in local richness of species before and after the K/Pg frontier was
broken by an abrupt increase of two to three times in number of
species.
After a time lapse of the order of
ten million years subsequent to a mass extinction, the original level of
biodiversity is restored in communities as a result of increased
speciation rate. Speciation rate apparently culminates roughly ten
million years after the end of a mass extinction event and remains
accelerated.
Discussion
Plant production increase has always
been linked to local biodiversity gain. In the last decade, there
were published papers which were against various conservationist
assumptions. Recent analyses refute the link between diversity loss
of plant species and ecosystem function, and question alleged
motivations of current biodiversity conservationism when confronted
with experimental results.
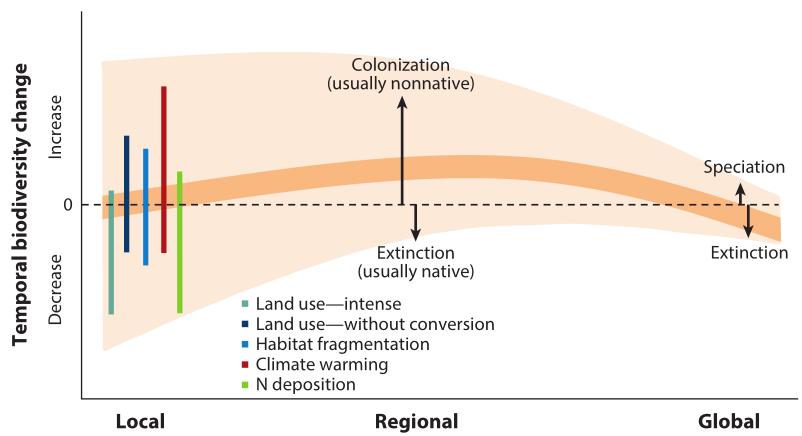
In the absence of total habitat
conversion, e.g., when a tropical forest becomes a parking lot or a
monoculture crop, local plant diversity declined very little or not
at all, on average, in the last century (figure 1, upper panel in B
and lower panel), nor there is available data that can forecast
otherwise in the future. Observed species diversity rates are
irrelevant in ecosystem function assessment, e.g., production and
nutrient cycling.
Species play more influential roles in
the local scale interface. There are as many increases as
reductions in biodiversity, especially of plant species in all scales
except global, which arguably registers decline.
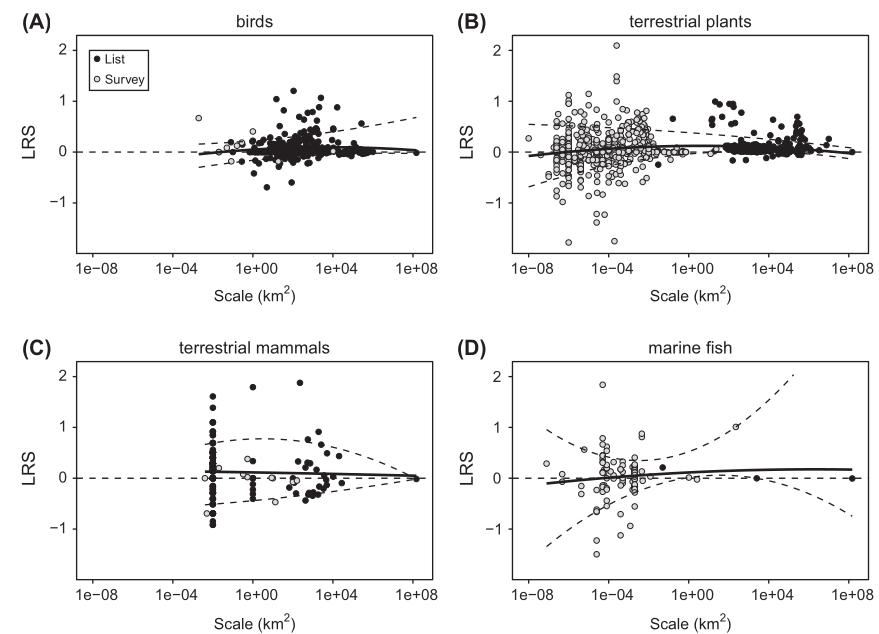
Figure
1.Upper panel:
these figures show a plot of LRS (log ratio of species richness)
versus log spatial scale, indicating trends (positive is an increase,
negative a decrease in richness). Light circles are survey-based
data. Dark circles are checklist-based data. Data are presented for
(A) terrestrial birds, (B) terrestrial plants, (C) terrestrial
mammals and (D) marine fishes during the Anthropocene. The solid line is
the best fit from a quadratic ordinary least squares regression and the
dashed lines are the 10 and 90% quantiles. Context dependence (scale)
allows a wide range of variation for each forcing in whatever
direction (Chase, 2019). Lower panel:
in comparison, similar graph for plants from Vellend (2017).
Conservation programmes decreed
“ecological successes” are opinions of each individual groups and
their goals debatable, e.g., how to reconcile conservation efforts
that require conflicting resources for various species (even with
human beings)? Hairy and feathery animals in the top of the food
chain are very much valued by people, however they occupy a more
plastic niche (place) in the ecosystem and are less important than
animals from the base of the food chain, such as cockroaches, which
do not attract as much attention.
Researchers from Rio de Janeiro
evaluated restoration plans of hundreds of medium and large-sized
localities in South Bahia in which there were vast Eucalyptus
forests. Those areas were compared to other tropical restoration
areas with regeneration time from half to 200 years. The response of
biodiversity ratio in naturally regenerated forests was more similar
to reference forests from countries with low, high, and very high
Human Development Index. This potentially reflects biodiversity
restoration projected by the Environmental Kuznets Curve (EKC).
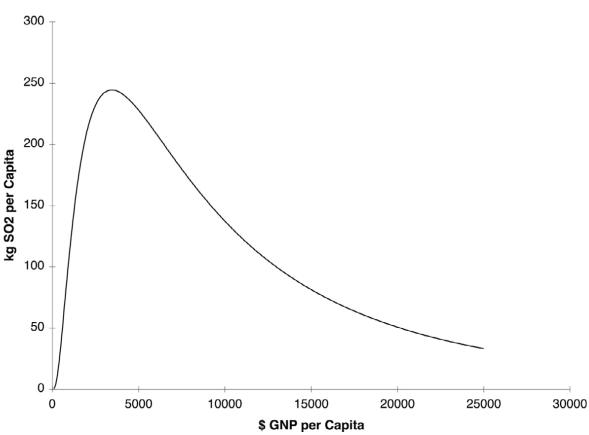
The EKC is used by economists for
projecting pollution emission vs per capita income. In early stages of
economic growth, human impacts increase, including pollution
emission, however after reaching certain development levels this
trend reverts. When the abscissa axis contains time and the ordinate
axis the level of some pollutant, it results in an
“inverted-U” curve of the pollutant level over time, which is
characteristic of EKC.
RED LISTS, INTERNATIONAL REPORTS FROM
IPBES AND LPI, BRAZILIAN MMA, ICMBIO, AND IBGE AGENCIES
In primary analysis, some weak points
can be verified about WWF Living Planet Index (LPI). In one index
from the report, the region with the highest ecological footprint is the
Neartic (includes the United States), whereas the Neotropical region has
a low ecological footprint. However, the Neartic region, within the
confidence levels, practically restores its LPI index to levels close
to 1970 marks, whereas the Neotropical region is presented as the
worst contributor for the decline in the global index throughout the
period of study. In order to conciliate these results, we must resort
to EKC.
There was debate in the scientific
community about WWF LPI data and assertions which stated that more
than 50% of animal species had gone extinct in the world. Such an
assertion turns out to be an artefact because less than 3% of the
index vertebrate population is distorting it. When these extreme decline
populations are taken out from the calculation, the global trend
turns to increase of species.
There are a couple authoritative
lists that register extinct and extinction risk species names. These
lists are used world-wide and are frequently cited reciprocally,
however their methodologies are strikingly different. In certain
lists, species that were not even properly described or seen only
once are considered extinct. Methodological changes that
hasten the verdict of extinction of species are problematic,
for example, mammals are frequently rediscovered, such an effect
is called the “Lazarus effect”.
In 1920, Arrhenius postulated that there
is a positive relation between area and number of species.
Wilson & MacArthur formulated the theory of island biogeography
in 1967, and Wilson based his allegations between species vs area.
However, this relation was put upside down and it was proposed that
habitat reduction must change species which in turn supposedly means
reduction in the number of population or disappearance of some
species, birds and great mammals being more susceptible to
extinction. Even though plausible, this postulate became one more
artifact amongst modern biologists.
Postulates of more than 27,000
extinctions within the period called by some Anthropocene, the
Intergovernmental Science-Policy Platform on Biodiversity and
Ecosystem Services (IPBES) and United Nations proved to be extremely
exaggerated.
Species subject to colonise only a
small area are really not deemed to have a great future, this being
the case of most extinct species of the IUCN Red List in the global
level, represented mainly by insect and insular species. When
European predators (such as rats, snakes, dogs, and cats) run across
new environments native species died, with 95% of mammal extinctions
being from insular and Australian species. The arrival of Europeans
to Australia, which had been isolated from the continent
for the previous 40 million years, was a single event.
Extinction lists from the IUCN Red
List, ordinances from Brazilian Ministry of the Environment nº 444 e
445 (17/Dec/2014), and lists produced by the Chico Mendes Institute
for Biodiversity Conservation (ICMBio) were analysed in the present
paper. Lists from these governmental entities were subsequently used
by the Brazilian Institute of Geography and Statistics (IBGE) in its
Ecosystem Accounts from 2020.
Table
1.Number of category changes that occurred between
2007 and 2021, as well as the proportion of criteria with non-genuine
(when new data, taxonomic revision and others become available),
genuine and hybrid motivations of the IUCN Red List.
Motivation
Changes
Proportion
Improved
Worsened
Same
Non-genuine
8288
88.31%
3756
4474
58
Genuine
1092
11.64%
163
921
8
Hybrid
5
0.05%
5
0
0
Total
9385
100.00%
3924
5395
66
The IUCN Red List shows 20 species as
extinct in Brazil in its Dec/2021 update. Regionally extinct species,
such as sharks, may migrate territory without necessarily damaging
ecological function. In the Red List, five plant species were
observed extinct, all poorly collected and restricted to relatively
small patches of Rio de Janeiro state, however their biological
families are far from danger (Myrtaceae and Sapotaceae).
Still, more recent studies made progress in the ecology of various
animal species, such as the scarab Megadytes ducalis, which
may aid in its rediscovery.
On the other hand, Brazilian IBGE
Ecosystem Accounts (2020) and ICMBio Red Book (2018) present ten
extinct animal species in Brazil. From those, five were regionally
extirpated (RE) and have got recovery potential in other lands or
countries around the world, and therefore ought not to be considered
extinct in the global scale.
The only tetrapods that can be
considered really extinct (EX) are Noronhomys vespuccii
(insular rodent from Fernando de Noronha) and Phrynomedusa
fimbriata (frog natural of Alto da Serra, Paranapiacaba,
Santo André-SP). Even with 10% left of the Atlantic Rainforest, only
the species Pauxi mitu (sin. Mitu mitu, Alagoas
helmeted curassow) is extinct in the wild (EW), and maybe Glaucidium
mooreorum (Pernambuco pygmy owl, still listed as critical risk
[CR] in the IUCN Red List), both restricted to central Pernambuco
state, the latter species discovered in an area of less than 5 Km²
and without vocalization records since 2001.
For their turn, Cichlocolaptes
mazarbarnetti (Atlantic rainforest Cryptic treehunter, last seen
in 2005) and Philydor novaesi (Alagoas foliage-gleaner,
considered rare, observed in secondary and selectively logged
forests, last seen in 2011) had their category worsened to EX by
classification methodologies adopted recently. See Table 1 for
motivations of category changes of IUCN Red List species from 2007 to
2021.
Conclusion
The EKC has received criticism since
its inception, partly because of differences in opinions about what
pollution is. Another problematic critique revolves about the duration of
time lapses needed to observe the effect. In general, EKC reveals
that human development sets excessive pollution in check over time
and restores environmental levels.
Scrutiny of extinction lists uncovers
important nuances for their proper interpretation. In multiple cases
their results and deductions are exaggerated. It is noted that the IUCN
Red List is more dependable than the IPBES report because the former uses
a more conservative methodology that only counts species with Latin
names, in contrast to IPBES which even considers species that have
not got a name. Recent methodology changes made many species to be
considered extinct. Brazilian lists also present exaggerated
diagnosis, disputably there being only two bird extinctions from
Atlantic rainforest. The species vs area relation is not kept when
there is habitat reduction.
Conservation efforts help to
alleviate or even revert extinctions. As long as species are not
insular nor inhabitant of restricted places and not subject to over
hunting by invading predator species, such as men, habitat reduction
is not an extinction sentence.
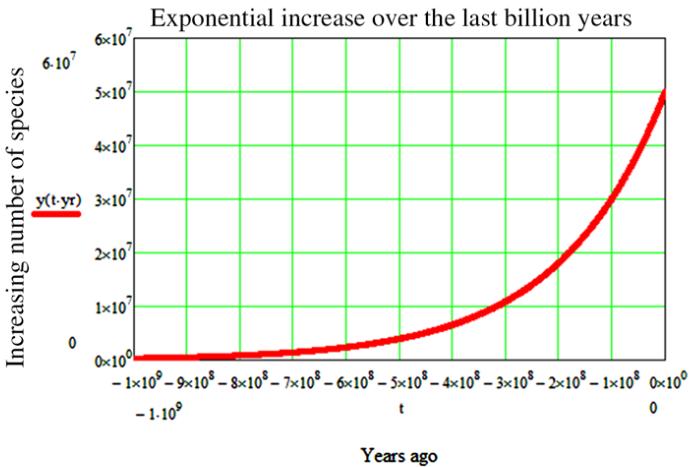
Figure
2. Since its inception, life on Earth
follows an exponential curve representing the growing number of
species (the graph does not take under account the well-known mass
extinctions) (Maccone, 2013).
Environmental
pressures benefit some species and harm others and biodiversity did
not decline in all levels in all places, arguably even increased in
the global level when a small percentage of extreme decline
populations (mainly from islands) is taken out from calculations.
Ecosystem function do not seem to be
hurt, in general, by
the observed rapid biodiversity compositional change,
especially in the plant and marine domains, even though species
homogenisation may be present in the regional and local scales.
Lastly, biodiversity generation
follows the first and second laws of thermodynamics for it improves
entropy generation and, therefore, increases entropy of the whole
system. There is no way we can brake this great biological reactor
that is Planet Earth (figure 2)!
Com maior tempo de evolução das espécies, menos mudanças podem
ser observadas, pois mutações geradas muito rapidamente por
pressões ambientais variantes não são possíveis de serem
percebidas com baixa resolução temporal, como nos estudos de
evolução fossilífera. Alterações ambientais promovem a “Corrida
da Rainha Vermelha” em diversos níveis de função e os organismos
sempre mudam mais ainda que para retornar e acompanhar um estado
anterior.
Lamarck, Darwin e Wallace são gradualistas em suas teorias de
evolução. O geólogo Niels Eldredge em 1972, com suas observações
de fósseis, verificou que há longos e tediosos períodos sem
mudança na morfologia de espécies que chamou de “estase” (de
estático) e que os mesmos fatores ambientais dirigindo extinções
também são responsáveis por padrões de especiação. Com isso,
formulou a teoria do “equilíbrio pontuado”, quando novos padrões
de especiação furam longos períodos monótonos de estase
biológica. Elementos genéticos transponíveis (transposons) são
essenciais nesse fenômeno (Casanova, 2020).
A pressão ambiental deve atingir toda a espécia geograficamente e
as mudanças morfológicas somente ocorrerão depois da evolução
genética e quando comunidades reprodutoras isoladas não puderem
mais trocar informação com outras espécies relacionadas
(subespécies).
Nas anotações de Darwin quando passou pela América Latina, cerca
de 1832, Brasil e Argentina, questionou qual motivo de não verificar
alta diversidade de alguns gêneros em sua longa transecção. Se o
espaço fosse mais importante do que o tempo na história da vida,
poder-se-ia explicar a estabilidade nas espécies em um território
continental integrado como o nosso ao longo do tempo, em contraste
com espécies relacionadas entre si porém distintas morfologicamente
que observaria em arquipélagos.
2. DISCUSSÃO
A taxa de diversidade pode ser calculada pela diferença entre as
taxas de especiação e extinção ao longo do espaço geográfico
(macroecologia) ou ao longo do tempo (macroevolução). Desde
aproximadamente 1600, há mudanças bem registradas na composição
de espécies de vários lugares no mundo.
Pimm (2014) faz um apanhado do número geral de espécies. Sem contar
as archaeas, bactérias, fungos e vírus, estima-se que haja por
volta de 1,5-1,9 milhões de espécies descritas (ver também Mora,
2011), enquanto a grande maioria ainda não foi. O número total de
plantas terrestres deva ser maior que 450 mil. Estimativas de números
de animais vão de três a 11 milhões de espécies no planeta Terra,
sendo estimados entre cinco e seis milhões de espécies de insetos
somente, enquanto outros autores questionam a possibilidade de uma
estimativa plausível quando consideram as incertezas nos números de
insetos e fungos. Espécies marinhas são estimadas entre 700 mil e
2,2 milhões, com somente aproximadamente 226 mil descritas. A média
de período de existência de espécies de invertebrados e
vertebrados verificada é de um a dez milhões de anos (vide taxas de
extinção, tabelas 1 e 2).
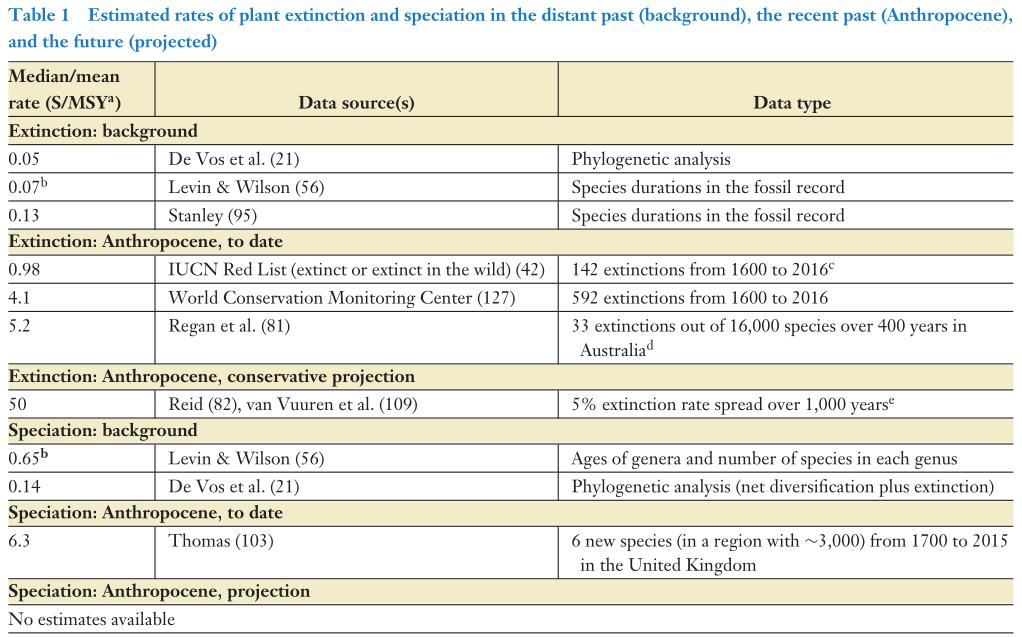
Tabela 1. Taxas
estimadas de extinção e especiação de plantas no passado distante
(fundo), passado recente (Antropoceno) e futuro (projetado). Espécies
por milhão de anos por ano ou, alternativamente, espécies por
espécies por milhão de anos; por exemplo, uma especiação estimada
em 1,0 S/MSY significa que a cada milhão de espécies uma nova
espécie surgirá a cada ano, ou, equivalentemente, cada espécie tem
a probabilidade de gerar uma espécie adicional a cada milhão de
anos (Vellend, 2017).
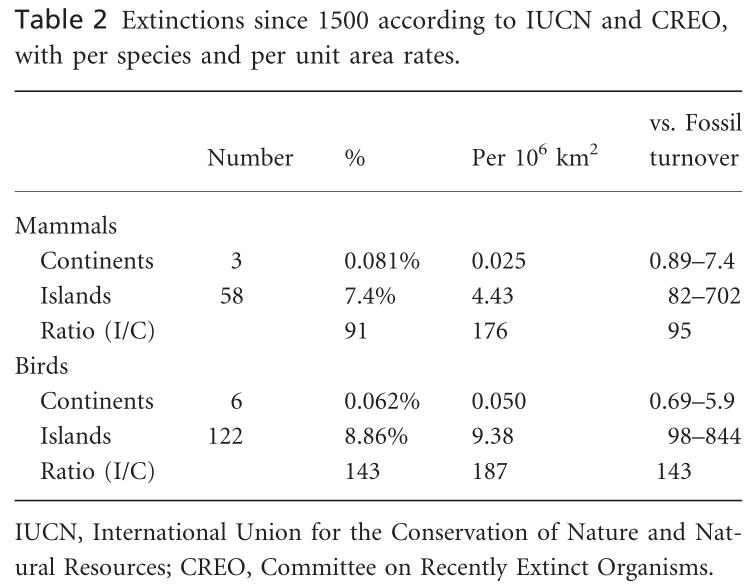
Tabela 2. Extinções
desde 1500 de acordo com o IUCN e CREO. Por base de unidade de área,
a taxa de extinção nas ilhas foi 177 vezes maior para os mamíferos
e 187 vezes maior para as aves do que nos continentes. A taxa
continental de extinção de mamíferos foi entre 0,89 e 7,4 vezes a
de fundo, enquanto a taxa de extinção de mamíferos de ilhas foi
entre 82 e 702 vezes a de fundo. A taxa de extinção de aves
continentais foi entre 0,69 e 5,9 vezes a de fundo, enquanto para
ilhas foi entre 98 e 844 vezes a de fundo. Razão das taxas de
ilha/continente (I/C). Extinções pré-históricas não
documentadas, principalmente em ilhas, ampliam essas tendências. As
taxas de extinção de ilhas são muito mais altas do que as taxas
continentais, em grande parte devido à introdução de predadores
alienígenas (incluindo o homem) e doenças (Loehle & Eschenbach,
2012).
Transições de espécie para espécie são difíceis de documentar.
Para ser demonstrar como uma espécie surgiu, gradualmente ou
repentinamente, são necessários estratos excepcionalmente
completos, com muitos animais mortos enterrados sob uma sedimentação
rápida e constante, o que não é comum em ambientes terrestres
(Gingerich, 1976, 1980).
Em um mundo físico heterogêneo e em mutação, nem tudo é um mar
de rosas para a biodiversidade. Variações espaçotemporais na
biodiversidade são a regra e não a exceção (Escarguel, 2011).
Na história geológica, houve cinco grandes eventos de extinção,
todos registrados no Fanerozoico, há 570 milhões de anos
e, na sua maioria, seus términos coincidiram com o início dos eventos de especiação.
O primeiro grande evento de especiação
foi a explosão Cambriana de animais marinhos esqueléticos
há aproximadamente 540 milhões de anos. O segundo foi o Grande
Evento de Biodiversificação do Ordoviciano (GOBE) (Webby, 2004).
Seria uma simplificação excessiva dizer que um evento dramático
desencadeou grandes pulsões de biodiversidade em todos os grupos
fósseis em nível global em um intervalo de tempo particular, por
exemplo, o GOBE não foi um evento único (Servais, 2021) (ver figura
1). Adendo: GOBE ocorreu
juntamente com o evento de extinção do Cambriano-Ordoviciano.
Close (2019) evidencia um aumento de diversidade na riqueza local
durante o Cretáceo (K) e Paleogêneo (Pg), seguido de relativa
estase até o presente nos tetrápodes terrestres. O padrão de
estase em riqueza local de espécies antes e depois da fronteira K/Pg
foi quebrado por um aumento abrupto de duas a três vezes no número
de espécies.
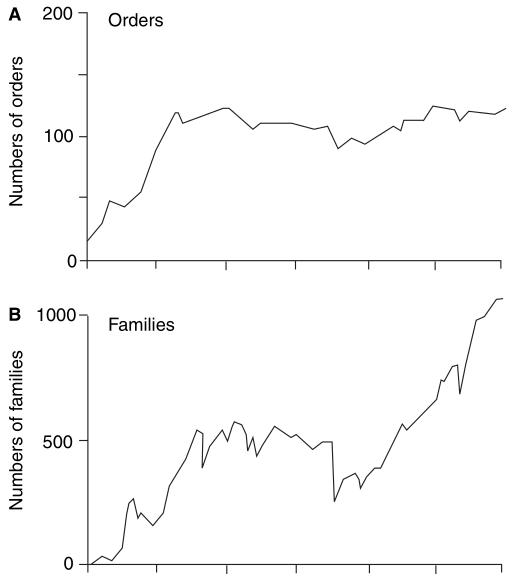
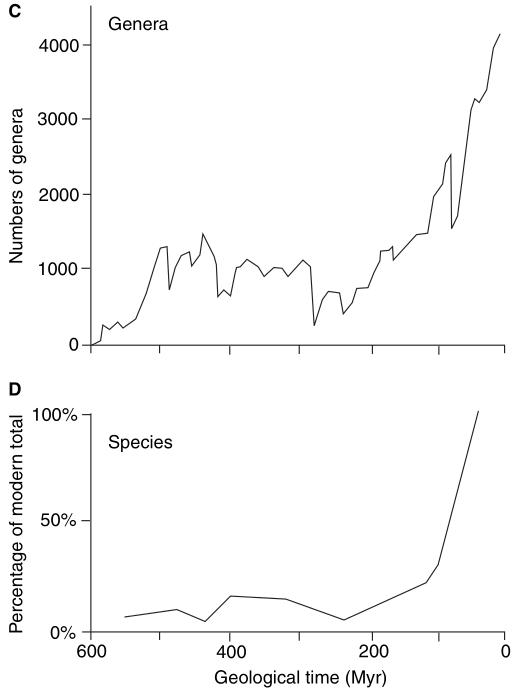
Figura 1. Padrões de
diversificação de animais marinhos com esqueletos bem formados,
contados como: A, ordens; B, famílias; C, gêneros; e D, espécies.
Contagens ordinal, familial e genérica são baseadas em dados
empíricos, enquanto que a curva de espécies é baseada em contagens
reais e em simulações. A curva é plotada como porcentagens do
total moderno (100%) (Benton & Emerson, 2007).
Após um espaço da ordem de dez milhões de anos, o nível de
biodiversidade original é restaurado nas comunidades como resultado
de uma maior taxa de especiação. A taxa de especiação
aparentemente culmina aproximadamente após dez milhões de anos
depois do final de uma extinção em massa (Flegr, 2009) e permanece
acelerada. A duração desse período não parece estar
correlacionado com o número de espécies (riqueza) afetadas pela
extinção (Kirchner, 2000).
Durante todo o período Fanerozoico, o número de eventos de
especiações foi quase igual com os de extinções, com uma
diferença de somente 0,1-1% (Close, 2019; Flegr, 2009).
Em outra frente, aumento de produtividade vegetal sempre foi
relacionado com aumento de diversidade local. Na última década,
apareceram artigos que vão na contramão de vários pressupostos
conservacionistas. Análise de Vellend (2013) contradiz o pressuposto
que diminuição da diversidade de espécies vegetais estaria ligada
a perda de função do ecossistema e questiona as motivações
alegadas para conservação da biodiversidade atual quando as põe em
cheque contra resultados experimentais.
Na ausência de conversão total de habitat, por exemplo uma floresta
tropical tornando-se um estacionamento ou uma plantação de
monocultura, a diversidade local de plantas declinou muito pouco ou
não declinou, em média, no último século (figura 2, painel
superior em B e painel inferior), nem há dados que indiquem que o
futuro será diferente. Diferenças observadas na taxa de diversidade
de espécies são fundamentalmente irrelevantes na avaliação de
função de ecossistema, como, por exemplo, produtividade ou ciclagem
de nutrientes.
As espécies exercem função de forma mais
influente na interface da escala local.
Há tantos aumentos quanto declínios de biodiversidade,
especialmente de espécies vegetais, em todas as escalas exceto na
global, que registra declínio.
Blowes (2019) mostra resultados de várias táxons (grupos de animais
marinhos e terrestres) em diferentes partes do mundo e verificou que
as tendências são indiscerníveis de zero (nota-se o erro
estatístico é equivalente ao sinal para parcelas de quase todas as
áreas de extensão, menos na maior), e que, em ambientes costeiros
marinhos, a biodiversidade tende a aumentar, no geral.
Baselga (2015) sugeriu que mudanças na cobertura da paisagem
agrícola tem pouco impacto na diversidade β temporal (turnover) das
assembleias (ou grupos) de aves continentais.
Em Daskalova e Dornelas (2020), no domínio terrestre, estudou-se a
prevalência de ganhos e perdas de cobertura de floresta e
verificou-se que mudança na floresta é um catalisador tanto para
maiores ganhos e perdas de biodiversidade e não somente um
catalisador para perdas.
Programas de conservação consagrados “sucessos ecológicos” são
opinião de cada grupo e seus propósitos debatíveis (Barnaud, 2021;
Cardinale, 2018), por exemplo como conciliar esforços de conservação
de espécies que necessitam de recursos conflitantes (até mesmo com
o ser humano) (Ceballos, 2005)?
Figura
2. Painel superior:
o LRS (razão logarítmica de riqueza de espécies) contra a escala
espacial logarítmica. Círculos claros são de dados provenientes de
observações e círculos escuros são baseadas em listas de
checagem. Dados apresentados para (A) aves terrestres, (B) plantas
terrestres, (C) mamíferos terrestres e (D) peixes marinhos durante o
Antropoceno. A linha sólida representa o melhor ajuste de uma
regressão quadrática ordinária dos mínimos quadrados e a linha
tracejada são os quantis de 10% e 90%. Dependência
de contexto (escala) cria uma ampla gama de variações para cada
forçante, em qualquer das direções (Chase, 2019). Painel inferior:
em comparação, gráfico similar para plantas terrestres de Vellend
(2017).
Pesquisadores do Rio de Janeiro (Latawiec, 2016) avaliaram o plano de
restauração de 284 localidades de tamanho médio e grandes na
Bahia, em que havia grandes plantações de Eucalyptus.
Foram comparadas com outras áreas de regeneração tropicais com
tempo de regeneração de meio a 200 anos. A resposta da proporção
da biodiversidade em florestas regeneradas naturalmente foi mais
similar em florestas de referência de países com valores de IDH
baixo, alto ou muito alto. Isso reflete, potencialmente, a
recuperação da biodiversidade prevista pela curva ambiental de
Kuznets.
A curva ambiental de Kuznets é usada por economistas para modelagem
de emissões de poluentes e renda per capta. Nos estágios iniciais
do crescimento econômico, impactos humanos aumentam, incluindo
emissões de poluentes, mas depois de certos níveis de
desenvolvimento, a tendência se reverte (figura 3) (Stern, 2004).
Figura 3. Curva
ambiental de Kuznets para emissões de dióxido de enxofre. Nota-se
que o impacto ambiental ou emissões per capta é uma curva em forma
de U invertido como uma função do PIB per capta (Stern, 2004).
2.1 LISTAS VERMELHAS E RELATÓRIOS
INTERNACIONAIS DO IPBES, LPI E NACIONAIS MMA, ICMBio E IBGE
Em análise, pude verificar alguns pontos fracos sobre o Living
Planet Index (LPI) da WWF. A pesquisa delimita o planeta em cinco
regiões. Em uma dos índices no relatório, a região com maior
pegada ecológica de consumo é justamente a neártica (inclui
Estados Unidos), já a região neotropical (inclui o Brasil) tem uma
pegada ecológica baixa. Porém, a região neártica, dentro dos
limites de confiança, praticamente restaura seu índice LPI a níveis
próximos de 1970, enquanto a região neotropical é apresentada como
a pior contribuidora para o declínio do índice global no período
estudado. Para conciliarmos esses resultados, devemos nos lembrar da
Teoria Ambiental de Kuznets, já estudada anteriormente.
Outros problemas com relação ao estudo da WWF sobre abundância de
espécies é a desproporção entre localidades de estudos no
hemisfério norte, além de poucos estudos no hemisfério sul, em
especial a região neotropical, acusada de ter perdido 89% da
abundância de espécies pelo relatório! As preocupações
levantadas implicam que o índice não é necessariamente um
indicador global, equilibrado e sem imparcialidade.
Ainda, utilizam na metodologia do LPI, programas de ciência do
cidadão, coletas feitas por cidadãos amadores e preocupante
tanto pela qualidade e uniformidade nos métodos e coleta quanto
resultados de identificações de espécies.
Leung (2020) reanalisou dados e afirmações do LPI da WWF, que dizia
que mais de 50% das espécies animais haviam sido extintas no mundo.
Observou-se que, em grande parte, esse dado é um artefato devido a
menos de 3% das populações de vertebrados do índice (maioria
insulares) estarem distorcendo o índice. Quando essas populações
em declínio extremo são excluídas, a tendência global muda para
aumento.
Há pelo menos duas listas oficiais autoritárias disponíveis que
contabilizam espécies com risco de extinção e extintas, como a Red
List da IUCN (International Union for Conservation of Nature and
Natural Resources) e do grupo Committe on Recently Extinct Organisms
(CREO), do Museus de História Natural de Nova York. A metodologia do
CREO é mais rígida. Por exemplo, primeiro deve ser demonstrado que
realmente há uma espécie envolvida para análise e é necessário
que haja taxonomia, espécimens e DNA (se possível). Nem todas
espécies da Red List apresentam dados completos sobre localização!
Algumas das restrições impedem que organismos avistados somente uma
vez em uma única ocasião possam ser considerados extintos, mas
ambas listas se citam reciprocamente.
O pesquisador climático independente Willis Eschenbach decidiu
verificar as duas listas e reavaliar as espécies de aves e animais.
Para reavaliar o postulado de Wilson e MacArthur (1967) na questão
específica sobre perda de habitat (e não sobre introdução de
espécies), Loehle e Eschenbach (2012) excluíram da análise todas
extinções de espécies insulares que tiveram contato com europeus
durantes explorações nos últimos 500 anos. Espécies introduzidas
em arquipélagos e na Austrália causaram verdadeiro estrago. Por
outro lado e para grande surpresa, foi verificado somente nove
extinções (seis aves e três mamíferos, até 2012) em todos os
continentes do mundo como África, Europa, Ásia, América do Norte e
América do Sul, nos 500 anos antecedentes.
Os postulados de Wilson de que haveria tido 27.000 extinções nesse
período (que coincide com estimativas do Intergovernmental
Science-Policy Platform on Biodiversity and Ecosystem Services
(IPBES) e Nações Unidas e de que as aves e os animais seriam os
mais afetados se mostraram extremamente exagerados.
Espécies sujeitas a colonizar somente uma pequena área realmente
não estão destinadas a ter um ótimo futuro. Esse é o caso da
grande maioria das espécies extintas da Red List. Conclui-se que
quando predadores europeus encontraram novos ambientes (ratos,
serpentes, cachorros e gatos), espécies nativas morreram, sendo que
95% das extinções de mamíferos são espécies insulares e
australianas. A chegada dos europeus na Austrália, que estava
isolada do continente por 40 milhões de anos anteriores, foi um
evento único.
O geólogo veterano e autor do livro “Inconvenient Facts: The
Science that Al Gore Doesn't Want You to Know”, Gregory Wrightstone
revelou em blogs (2019a e 2019b) uma análise aprofundada da Red List
e uma interpretação detalhada bem diferente do encontrado pelo
IPBES, expondo resultados parecidos com Loehle e Eschenbach (2012).
Ainda, Fisher e Blomberg (2011) mostraram que espécies de mamíferos
são frequentemente redescobertas, chamado “efeito Lázaro”.
Analisei as listas de extinção da Red List, as portarias do
Ministério do Meio Ambiente nº 444 e 445 (17/12/2014) e a Lista
Vermelha produzida pelo Instituto Chico Mendes de Conservação
(ICMBio), quais listas foram por sua vez utilizadas pelo Instituto
Brasileiro de Geografia e Estatística (IBGE) em suas Contas de
Ecossistemas de 2020.
Motivações
Mudanças
Proporção
Melhorou
Piorou
Igual
Não-genuínas
8288
88,31%
3756
4474
58
Genuínas
1092
11,64%
163
921
8
Híbridas
5
0,05%
5
0
0
Total
9385
100,00%
3924
5395
66
Tabela 3. Número
mudanças de categoria que ocorreram de 2007 a 2021 e proporção da
utilização de critérios com motivações não-genuínas (quando
novos dados são avaliados, revisão taxonômica, etc), genuínas e
híbridas da Red List da IUCN (arquivo pessoal).
A Red List aponta 20 espécies como extintas no Brasil em sua última
atualização (Dez/2021). Espécies extintas regionalmente, como
tubarões, podem migrar de território, como veremos, sem
necessariamente prejuízo de função de ecossistema. Na Red List,
observa-se cinco espécies vegetais extintas, todas restritas a
relativamente pequenas áreas do estado do Rio de Janeiro e pouco
coletadas, mas suas famílias vegetais estão longe de correr perigo
(Myrtaceae e Sapotaceae). Ainda, estudos recentes estão
possibilitando melhor conhecimento da ecologia de várias espécies
de animais, como escaravelhos Megadytes ducalis, o que pode
ajudar em seu redescobrimento (por exemplo, Hendrich, 2019).
Já as Contas de Ecossistema (2020) do IBGE e Livro Vermelho do
ICMBio (2018), apresentam dez espécies de animais extintos no
Brasil. Deles, cinco foram extirpados regionalmente (RE) e apresentam
potencial de recuperação em outros países ou áreas, e portanto
não podem ser considerados extintos em escala global.
Os únicos tetrápodes que podem ser consideradas realmente extintos
(EX) são Noronhomys vespuccii (roedor insular) e Phrynomedusa
fimbriata (perereca oriunda unicamente de Alto da Serra,
Paranapiacaba, Santo André/SP). Mesmo com 10% restante de mata
atlântica, somente a espécie Pauxi mitu (sin. Mitu mitu)
é tida como Extinta na Natureza (EW), e talvez Glaucidium
mooreorum (na Red List ainda consta como Criticamente em Perigo
[CR]), ambas restritas ao centro de Pernambuco, esta última
descoberta em uma área de menos de 5 km^2 e sem vocalização
gravada desde 2001.
Já Cichlocolaptes mazarbarnetti (ave da mata atlântica
avistada pela última vez em 2005) e Philydor novaesi
(considerada rara, observada em florestas secundárias e
desflorestadas seletivamente, observada pela última vez em 2011)
pioraram de categoria para EX com novas metodologias de classificação
adotadas recentemente. Ver, por exemplo, a tabela 3 para as
motivações de mudanças de categoria entre 2007 e 2021.
3. CONCLUSÃO
A
curva ambiental de Kuznets tem recebido críticas desde sua
formulação. Isso se deve, em parte, por diferenças de opiniões
sobre o que seria poluição. Por exemplo, emissões de CO2tem
aumentado continuamente nas últimas décadas, porém, sabemos que
CO2nãoé,
na realidade, um poluente (Zhu, 2016). Outro problema nas críticas é
que certos bancos de dados utilizados são de análises de curta
duração temporal. De forma geral, a curva de Kuznets revela que o
desenvolvimento humano barra a poluição excessiva ao longo do tempo
e restaura níveis ambientais.
Análises das listas de extinção mostraram nuances importantes para
correta interpretação pois em muitos casos as análises e
conclusões são exageradas. Nota-se que a Red List é mais confiável
que o relatório do IPBES pois utiliza metodologia mais precisa e
melhor definida, contando espécies com nomes em Latim ao contrário
do IPBES que calcula espécies que sequer têm nome, porém mudanças
recentes de metodologia fizeram com que várias espécies sejam
consideradas extintas. As listas brasileiras também apresentam
diagnósticos exagerados, disputavelmente havendo extinção de
somente duas aves da mata atlântica em análise aprofundada.
Esforços de conservação ajudam a mitigar e até mesmo reverter
extinções (Pereira, 2012). Contanto
que não sejam espécies insulares ou que habitem lugares restritos,
e que não sejam caçadas até a morte por espécies predadoras
invasoras, como o homem, a redução de habitat por si não é uma
sentença de extinção.
Figura 4. Desde seu
início, a vida na Terra segue uma curva exponencial representativa
do crescente número de espécies (o gráfico não leva em
consideração as bem conhecidas extinções em massa) (Maccone,
2013).
Pressões ambientais beneficiam algumas
espécies e prejudicam outras e a biodiversidade não declinou em
todos os níveis ecológicos ou em todos os lugares. A função dos
ecossistemas não parece estar prejudicada, de maneira geral, pela
rápida mudança de composição observada, especialmente nos
domínios vegetal e marinho, em que pese haver uma homogenização de
espécies nas escalas local e regional.
Por fim, a geração de
biodiversidade obedece à segunda lei da termodinâmica pois aumenta
a geração de entropia, e portanto a entropia total do sistema
(Skene, 2005). Não há como frearmos esse grande reator biológico
que é o planeta Terra (figura 4)!
BIBLIOGRAPHICAL CITATIONS
REFERÊNCIAS BIBLIOGRÁFICAS
BARNAUD, C. et al. Is forest regeneration good for biodiversity?
Exploring the social dimensions of an apparently ecological debate.
Environmental Science & Policy, v. 120, p. 63–72, jun.
2021.
BASELGA, A.; BONTHOUX, S.; BALENT, G. Temporal Beta Diversity of Bird
Assemblages in Agricultural Landscapes: Land Cover Change vs.
Stochastic Processes. PLOS ONE, v. 10, n. 5, p. e0127913, 26
maio 2015.
BENTON, M. J.; EMERSON, B. C. HOW DID LIFE BECOME SO DIVERSE? THE
DYNAMICS OF DIVERSIFICATION ACCORDING TO THE FOSSIL RECORD AND
MOLECULAR PHYLOGENETICS. Palaeontology, v. 50, n. 1, p. 23–40,
jan. 2007.
BLOWES, S. A. et al. The geography of biodiversity change in marine
and terrestrial assemblages. Science, v. 366, n. 6463, p.
339–345, 18 out. 2019.
CASANOVA, E. L.; KONKEL, M. K. The Developmental Gene Hypothesis for
Punctuated Equilibrium: Combined Roles of Developmental Regulatory
Genes and Transposable Elements. BioEssays, v. 42, n. 2, p.
1900173, fev. 2020.
CEBALLOS, G. et al. Global Mammal Conservation: What Must We Manage?
Science, v. 309, n. 5734, p. 603–607, 22 jul. 2005.
CHASE, J. M. et al. Species richness change across spatial scales.
Oikos, v. 128, n. 8, p. 1079–1091, ago. 2019.
CLOSE, R. A. et al. Diversity dynamics of Phanerozoic terrestrial
tetrapods at the local-community scale. Nature Ecology &
Evolution, v. 3, n. 4, p. 590–597, abr. 2019.
DASKALOVA, G. N. et al. Landscape-scale forest loss as a catalyst of
population and biodiversity change. p. 8, 2020.
ESCARGUEL, G. et al. Biodiversity is not (and never has been) a bed
of roses! Comptes Rendus Biologies, v. 334, n. 5–6, p.
351–359, maio 2011.
FISHER, D. O.; BLOMBERG, S. P. Correlates of rediscovery and the
detectability of extinction in mammals. Proceedings of the Royal
Society B: Biological Sciences, v. 278, n. 1708, p. 1090–1097,
7 abr. 2011.
GINGERICH, P. D. Evolutionary Patterns in Early Cenozoic Mammals.
Annual Review of Earth and Planetary Sciences, v. 8, n. 1, p.
407–424, maio 1980.
GINGERICH, P. D. Paleontology and Phylogeny: Patterns of Evolution at
the Species Level in Early Tertiary Mamals. American Journal of
Science, v. 276, p. 1-28, jan. 1976.
HENDRICH, L.; MANUEL, M.; BALKE, M. The return of the Duke—locality
data for Megadytes ducalis Sharp, 1882, the world’s largest diving
beetle, with notes on related species (Coleoptera: Dytiscidae).
Zootaxa, v. 4586, n. 3, p. 517, 18 abr. 2019.
KIRCHNER, J. W.; WEIL, A. Delayed biological recovery from
extinctions throughout the fossil record. Nature, v. 404, n.
6774, p. 177–180, mar. 2000.
LATAWIEC, A. E. et al. Natural regeneration and biodiversity: a
global meta-analysis and implications for spatial planning.
Biotropica, v. 48, n. 6, p. 844–855, nov. 2016.
LEUNG, B. et al. Clustered versus catastrophic global vertebrate
declines. Nature, v. 588, n. 7837, p. 267–271, 10 dez. 2020.
LOEHLE, C.; ESCHENBACH, W. Historical bird and terrestrial mammal
extinction rates and causes: Bird and mammal extinction rates and
causes. Diversity and Distributions, v. 18, n. 1, p. 84–91,
jan. 2012.
MACARTHUR, R. H.; WILSON, E. O. The Theory of Island Biogeography.
Princeton University Press, p. 1-203. 1967.
MACCONE, C. SETI, Evolution and Human History Merged into a
Mathematical Model. International Journal of Astrobiology, v.
12, n. 3, p. 218–245, jul. 2013.
MORA, C. et al. How Many Species Are There on Earth and in the Ocean?
PLoS Biology, v. 9, n. 8, p. e1001127, 23 ago. 2011.
PEREIRA, H. M.; NAVARRO, L. M.; MARTINS, I. S. Global Biodiversity
Change: The Bad, the Good, and the Unknown. Annual Review of
Environment and Resources, v. 37, n. 1, p. 25–50, 21 nov. 2012.
PIMM, S. L. et al. The biodiversity of species and their rates of
extinction, distribution, and protection. Science, v. 344, n.
6187, p. 1246752, 30 maio 2014.
SERVAIS, T.; CASCALES-MIÑANA, B.; HARPER, D. A. T. The Great
Ordovician Biodiversification Event (GOBE) is Not a Single Event.
Paleontological Research, v. 25, n. 4, 1 out. 2021.
SKENE, K. Life’s a Gas: A Thermodynamic Theory of Biological
Evolution. Entropy, v. 17, n. 12, p. 5522–5548, 31 jul.
2015.
STERN, D. I. The Rise and Fall of the Environmental Kuznets Curve.
World Development, v. 32, n. 8, p. 1419–1439, ago. 2004.
VELLEND, M. et al. Global meta-analysis reveals no net change in
local-scale plant biodiversity over time. Proceedings of the
National Academy of Sciences, v. 110, n. 48, p. 19456–19459, 26
nov. 2013.
VELLEND, M. et al. Plant Biodiversity Change Across Scales During the
Anthropocene. Annual Review of Plant Biology, v. 68, n. 1, p.
563–586, 28 abr. 2017.
WEBBY, B. et al. (EDS.). The Great Ordovician Biodiversification
Event Columbia University Press, 2004.